Les échelles de la biodiversité


Les échelles de la biodiversité
La biodiversité est un concept des années 1980 qui se définit comme la diversité du vivant et sa dynamique. Diversité du vivant = ensemble d’espèces en interaction entre-elles et avec le milieu. Il existe différentes échelles de cette biodiversité.
I. Échelle des espèces
Au sein d’un écosystème forestier, il y a un biotope et une biocénose :
– Le biotope est le milieu et l’ensemble des paramètres physiques et chimiques de ce milieu : la température, le degré d’hydrométrie, l’intensité lumineuse, etc.
– La biocénose est l’ensemble des espèces qui évoluent en interaction entre-elles.
Dans un écosystème forestier extrêmement simplifié, ces espèces établissent des relations trophiques « qui mange qui » = notion de réseau trophique ou de chaîne alimentaire. Dans un réseau trophique, on représente une flèche de l’individu qui mange vers l’individu qui est mangé.
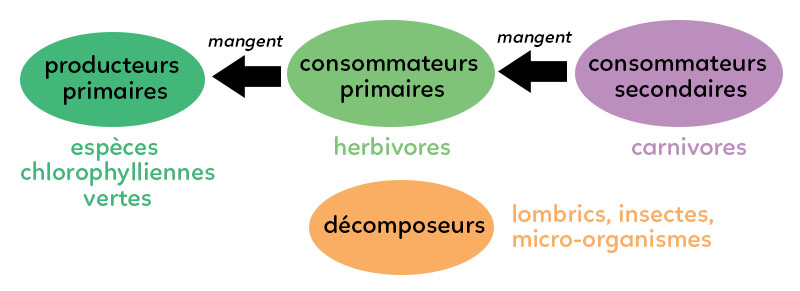
A. Les producteurs primaires
Au sein de l’écosystème forestier, on trouve les espèces chlorophylliennes vertes qui réalisent la photosynthèse, et qui, par le biais de l’énergie lumineuse (hµ) transforment la matière minérale en matière organique. Ceux sont des producteurs primaires de matière organique par autotrophie. Ces producteurs primaires riches en chlorophylle font entrer le carbone minéral et l’assimilent en carbone organique. Les arbres, comme le chêne, les arbustes ou les fougères forment ainsi le groupe des producteurs primaires.
B. Les consommateurs primaires
Ces producteurs primaires sont ensuite consommés par des consommateurs primaires : des herbivores. Les petits oiseaux tels que les mésanges ou le lapin (qui mange les racines) sont des consommateurs primaires. Mais également, les insectes comme les coléoptères qui consomment la partie chlorophyllienne des producteurs primaires.
C. Les consommateurs secondaires
Ces consommateurs primaires sont à leur tour mangés par un consommateur secondaire. Le consommateur secondaire peut être, par exemple, une buse (rapace) qui mange la mésange (qui elle-même mange une partie du chêne). Il y a aussi le lapin qui est mangé par le renard. Le renard et la buse sont des carnivores, des consommateurs secondaires.
D. Les décomposeurs
Dans cet écosystème forestier, les décomposeurs interviennent plutôt dans la partie du sol et favorisent le processus d’humidification. Dans les décomposeurs, il y a les lombrics (vers de terre), un certain nombre d’insectes et les micro-organismes comme les bactéries qui participent à la décomposition de la matière organique morte.
II. Échelle des populations
À l’échelle de l’écosystème, on a vu une illustration de la biodiversité avec de nombreuses espèces différentes. On peut passer à l’échelle en dessous, celle de la population pour illustrer la biodiversité.
Dans cet exemple d’écosystème forestier, une population serait un ensemble d’individus appartenant à la même espèce ayant autant de probabilité de se rencontrer en termes de reproduction. Par exemple, les mésanges au sein d’un chêne forment une population. D’un point de vue dynamique, évolutif, on peut la comparer à une autre population de mésanges. En effet, des études faites en Angleterre sur des mésanges qui ont acquis la capacité d’ouvrir des bouteilles de lait montrent qu’il y a un nouveau caractère que l’on va découvrir chez cette population-là. On est dans un cas de biodiversité, non plus à l’échelle de l’écosystème mais à l’échelle des populations. On va pouvoir comparer des populations et comprendre les mécanismes évolutifs différents.
II. Échelle des individus, échelle cellulaire, échelle moléculaire
On peut comprendre la biodiversité à l’échelle individuelle, à l’échelle de l’individu. Ainsi, on peut comparer une mésange précise (par exemple morphologiquement par le phénotype macroscopique qui est l’ensemble des caractères que l’on observe à l’œil nu) à une autre.
On peut continuer à descendre dans les échelles d’observation, pour montrer une certaine biodiversité inter-individuelle à l’échelle cellulaire voire à l’échelle moléculaire, en comparant non pas les gènes mais les allèles. En effet, montrer une diversité allélique, c’est montrer une biodiversité à l’échelle de l’individu.
La notion d'espèce
La notion d’espèce est un concept compliqué qui se définit par plusieurs approches.
I. Définition biologique
La définition la plus communément admise est celle d’Ernst Mayr qui est une définition biologique. Il dit que deux individus appartiennent à la même espèce, si et seulement si, ces deux individus sont interféconds et que leur descendance est fertile, c’est-à-dire qu’à leur tour les descendants pourront avoir une descendance.
Par exemple avec l’espèce cheval et l’espèce âne : la descendance obtenue entre un âne mâle et une jument est un individu (donc on peut supposer que l’interfécondité fonctionne et est réelle) qui s’appelle une mule ou un mulet. Cet individu est stérile, non fertile.
Donc cette définition fonctionne bien, mais elle a des limites. Par exemple, il existe l’espèce chien et l’espèce loup, lorsqu’il y a reproduction entre les deux, cela donne un nouvel individu (chien-loup ou crocotte) fertile. Ainsi, cette définition sur les hybrides trouve des limites.
De même, lorsqu’on travaille sur deux petites espèces d’insecte, que l’on voit deux individus, on se pose la question en laboratoire de savoir s’ils appartiennent à la même espèce ou non. On peut alors espérer que la fécondation ait lieu entre ces deux individus, mais en laboratoire ce n’est pas toujours évident d’obtenir une reproduction. Là encore, on est face à la limite de cette définition biologique.
Enfin, quand on pense aux espèces fossiles, par exemple les dinosaures, on a de nouveau les limites de cette définition. Comment savoir que deux dinosaures appartiennent à des espèces différentes ou non, puisqu’on n’a pas la connaissance de savoir s’ils pouvaient se reproduire entre-eux (car ils n’existent plus) ?
Il existe donc d’autres définitions de la notion d’espèce.
II. Définition génétique
Cette définition se base sur le caryotype qui est une représentation des chromosomes par couple d’homologues du plus grand au plus petit. Par exemple, dans l’espèce humaine, nous avons 46 chromosomes : ceci est un de nos attributs. Si l’on prend l’espèce chimpanzé, il y a 48 chromosomes. On a donc deux caryotypes différents, donc deux espèces différentes.
Parfois, il existe des caryotypes identiques (même nombre de chromosomes) mais pourtant des espèces différentes. Il faut alors se placer à l’échelle du génome (l’ensemble des gènes au sein d’une espèce). Dans l’espèce humaine, nous sommes à 22 000 ou 23 000 gènes et beaucoup plus d’allèles (qui sont les versions d’un gène). Ainsi, pour comparer des individus (savoir s’ils appartiennent ou non à la même espèce), on peut travailler sur des génomes du séquençage d’ADN et conclure à l’appartenance ou non des individus à la même espèce.
III. Définition phénotypique
Phéno : ce qui est visible, ce que l’on peut observer à différentes échelles : avec les yeux (échelle macroscopique), avec un microscope (échelle microscopique), avec des outils pour descendre à l’échelle moléculaire (échelle nanoscopique).
C’est donc travailler sur des ressemblances anatomiques. On prend deux individus et l’on se demande s’ils appartiennent à la même espèce. S’ils se ressemblent, il est fort probable que oui. Mais il existe des limites.
Par exemple, au sein de l’espèce lion, si l’on prend le mâle et la femelle, ils ont un ensemble d’attributs phénotypiques différents et pourtant ils appartiennent à la même espèce : même caryotype, même génome.
Un autre exemple, dans un cas de dimorphisme sexuel, c’est-à-dire que le mâle et la femelle sont très différents, on peut penser aux insectes comme le lucane cerf-volant avec un mâle phénotypiquement très différent de la femelle. Si on regarde les poissons, on a un exemple de dimorphisme sexuel poussé à son extrême avec la caulophryne : la femelle est énorme alors que le mâle vit presque en parasite sur la femelle.
IV. Définition écologique
Dans cette définition, deux individus appartiennent à la même espèce s’ils occupent la même niche écologique. Ceci est pratique lorsqu’on se place à l’échelle de l’écosystème, quand on travaille sur le terrain, mais il y a aussi des limites.
Par exemple, au sein de l’espèce grenouille Reinette, l’adulte n’occupe pas la même niche écologique dans le cycle de développement que le têtard, ils n’ont pas la même alimentation, ils ne vivent pas exactement au même endroit.
Diversité au sein d'une espèce : diversité génétique, allélique (ABO)
Pour illustrer la biodiversité au sein d’une espèce, on travaille à l’échelle des gènes et plus précisément des allèles, puisque les différentes versions d’un gène s’appellent des allèles.
On remarque au sein de l’espèce humaine une diversité individuelle : dans le cas des groupes sanguins, il y a des individus du groupe A, d’autres du groupe B, d’autres du groupe AB et enfin du groupe O.
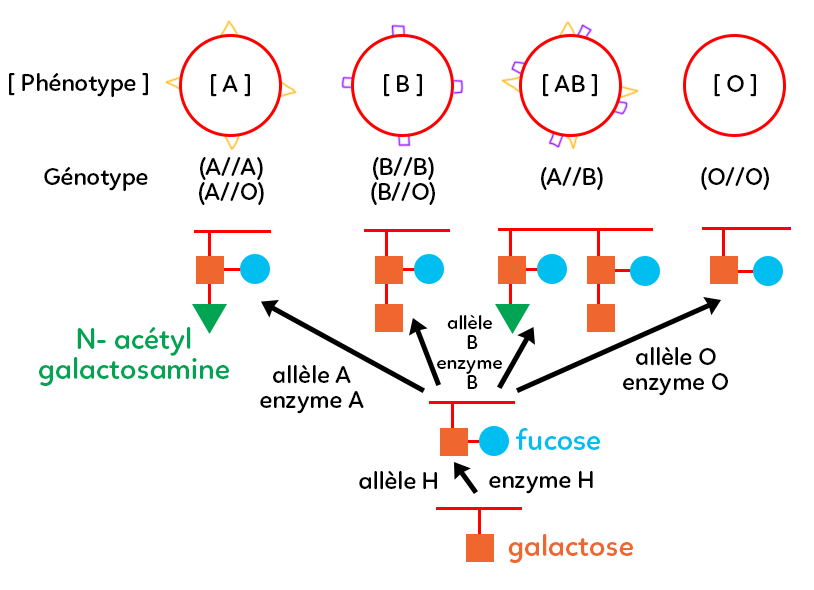
I. Les phénotypes (caractères observables)
– Pour le groupe sanguin A, on trouve microscopiquement et moléculairement des marqueurs moléculaires à la surface des globules rouges.
– Pour le groupe sanguin B, il y a des marqueurs moléculaires présents à la surface des globules rouges qui ne sont pas les mêmes que ceux du groupe sanguin A.
– Pour le groupe sanguin AB, on trouve à la surface des globules rouges à la fois les marqueurs du groupe A et les marqueurs du groupe B.
– Pour le groupe sanguin O, il n’y a pas de marqueurs moléculaires.
Le groupe sanguin AB peut recevoir du sang des individus du groupe A, du groupe B, du groupe AB et du groupe O. On dit que l’on est receveur universel. À l’inverse, pour le groupe sanguin O, on peut donner du sang à tous les groupes, donc on est donneur universel, mais on ne peut recevoir que du groupe O. Ces quatre phénotypes (A, B, AB, O) se représentent toujours entre crochets [ ].
II. Les génotypes
A. Marqueurs moléculaires : galactose et fucose
Sur le schéma, les traits rouges représentent la membrane du globule rouge. En bas du schéma, un sucre est accroché sur cette membrane : le galactose (carré orange). Un autre sucre, le fucose (rond bleu) va venir s’accrocher sur le galactose grâce à une protéine (enzyme). Cette enzyme s’appelle enzyme H car elle est codée par un gène, plus précisément une version allélique H. Lorsque l’on a le gène fucose, alors on a l’enzyme H. Ainsi, dans le génotype (l’ensemble des allèles) on a nécessairement l’allèle H.
B. Versions alléliques possibles
Sur ce support constitué d’un galactose et d’un fucose, au sein du chromosome 9 (groupe sanguin), on a un gène qui a plusieurs versions :
– La version allèlique A qui code pour une enzyme A. Cette enzyme vient accrocher sur le galactose un sucre qui s’appelle le N-acétylgalactosamine (triangle vert). On a donc un ensemble moléculaire carré/rond/triangle qui donne le marqueur moléculaire A. Une personne du groupe A a forcément le N-acétylgalactosamine accroché par l’enzyme A, codé par l’allèle A. Cet individu est donc nécessairement de génotype A (il a l’information A). Il peut être A//A (homozygote pour ce gène) ou A//O car l’allèle A est dominante sur l’allèle O.
– La version allélique B qui code pour une enzyme B. Le marqueur moléculaire carré/carré/rond est différent de celui du A, puisque l’on n’a pas la même enzyme. On a cette enzyme B car on a l’allèle B. Donc, ce phénotype B est permis par un génotype avec l’allèle B. Soit B//B, soit B//O.
– La version allélique AB : on a les marqueurs A et les marqueurs B, donc nécessairement on a l’enzyme B qui permet de fabriquer le marqueur carré/carré/rond mais aussi l’enzyme A. On a, à chaque fois, deux chromosomes, puisque l’humain est une espèce diploïde. Il y a un chromosome qui possède la version allélique A et l’autre qui possède la version allélique B. Le groupe sanguin AB est donc obligatoirement de génotype A//B. Puisque les deux s’expriment, on parle de codominance des allèles.
– La version allélique O n’a pas de marqueur moléculaire, il n’y a que carré/rond à la surface des globules rouges (galactose + fucose), car au niveau du chromosome 9, on a une version O. Quand on compare la longueur des nucléotides des différentes versions, pour l’allèle O, il y a un nucléotide en moins. Certainement une mutation par délétion. Ce nucléotide en moins implique que l’enzyme O ne fonctionne pas. Elle n’accroche rien sur ce motif-là. Autrement dit, à la surface des globules rouges, il y a seulement un galactose et un fucose. Ainsi, le génotype est nécessairement O//O, car O est un allèle récessif. Quand on est de phénotype O, on est nécessairement homozygote O//O.
Remarque
On peut être de phénotype O et pourtant posséder les allèles A, B, et AB car les groupes sanguins A, B et O dépendent des allèles A, B et O mais aussi, en amont, de l’allèle H.
Un individu qui présente une mutation de l’allèle H qui donnerait un allèle h, ne va pas pouvoir accrocher de fucose. Or, si l’on bloque la voie de biosynthèse (bas du schéma), bien que l’allèle A ou l’allèle B soit présente, on ne fabriquera rien car l’enzyme A ne fonctionne qu’avec le principe d’accroche avec le fucose. Ainsi, on peut être du groupe sanguin O avec les allèles A, B voir AB, si et seulement si, on a une enzyme H non fonctionnelle, c’est-à-dire de génotype h.