Méiose et répartition allélique pour 2 gènes indépendants
- 1
- 2Recherche du locus d'un gène
- 3Accidents méiotiques et évolution
- 4Étude de cas : risque de transmission de la myopathie de Duchenne
- 5Transferts horizontaux et endosymbiose
- 6Mutations, clones cellulaires et génome de l'individu
- 7Survie, reproduction et équilibre de Hardy-Weinberg
- 8Étude d'un cas de spéciation



Méiose et répartition allélique pour 2 gènes indépendants
Exercice : Méiose et répartition allélique pour 2 gènes indépendants
ÉNONCÉ
À partir de l’étude du document suivant, dresser un tableau de croisement (appelé aussi échiquier de croisement, ou tableau de gamètes) correspondant au deuxième croisement réalisé (F1 x P2) et permettant de comprendre la diversité des phénotypes obtenus en F2.
Puis schématiser les principales étapes de la répartition des chromosomes au cours de la méiose chez F1, expliquant les proportions obtenues en F2.
Pour simplifier on représentera la méiose d’une cellule avec 2n=4 chromosomes.
DOCUMENT
On étudie dans un laboratoire une espèce de monstres, dont le poil peut être roux ou bleuté. La tête peut également porter des cornes, ou pas.
On effectue un premier croisement d’un monstre P1 de souche pure de phénotype poil roux et avec des cornes avec un monstre P2 de souche pure de phénotype poil bleuté et sans corne. On obtient une première génération de monstres appelée F1, qui ont tous le poil roux et des cornes.
Le deuxième croisement (appelé TEST CROSS, ou croisement-test) entre des monstres de la F1 avec des monstres P2 donne les résultats suivants en F2 :
– 32 monstres à poil roux et avec cornes ;
– 34 monstres à poil roux et sans corne ;
– 31 monstres à poil bleuté et avec cornes ;
– 33 monstres à poil bleuté et sans corne.
Recherche du locus d'un gène
Exercice : Recherche du locus d’un gène
ÉNONCÉ
Utiliser des arguments du cours et du document afin de démontrer, par un raisonnement scientifique, sur quel chromosome se trouve le locus du gène aristaless.
Illustrer par un schéma simplifié le phénomène méiotique identifié, et qui permet ici de conclure.
DOCUMENT
La drosophile possède un caryotype diploïde constitué de 2n=8 chromosomes. On s’intéresse ici aux chromosomes non sexuels 2, 3 et 4, et on prend comme référence un gène situé sur chaque chromosome :
– Gène vestigial du chromosome 2 : donnant des ailes de longueur normale pour l’allèle vg+ (dominant) et des ailes raccourcies (vestigiales) pour l’allèle vg (récessif).
– Gène sépia du chromosome 3 : donnant des yeux rouges pour l’allèle se+ (dominant) et des yeux foncés (sépia) pour l’allèle se (récessif).
– Gène bent du chromosome 4 : donnant des ailes le long du corps pour l’allèle be+ (dominant) et des ailes écartées pour l’allèle be (récessif).
On cherche le locus du gène aristaless donnant des antennes de longueur normale pour l’allèle al+ (dominant) et des antennes courtes pour l’allèle al (récessif).
On croise des drosophiles hétérozygotes pour les 4 gènes étudiés ici avec des drosophiles homozygotes récessives pour ces mêmes gènes.
On obtient :
44 % de drosophiles aux ailes normales et antennes normales.
4 % de drosophiles aux ailes vestigiales et antennes normales.
6 % de drosophiles aux ailes normales et antennes courtes.
46 % de drosophiles aux ailes vestigiales et antennes courtes.
25 % de drosophiles aux yeux rouges et antennes normales.
25 % de drosophiles aux yeux sépia et antennes normales.
25 % de drosophiles aux yeux rouges et antennes courtes.
25 % de drosophiles aux yeux sépia et antennes courtes.
25 % de drosophiles aux ailes le long du corps et antennes normales.
25 % de drosophiles aux ailes écartées et antennes normales.
25 % de drosophiles aux ailes le long du corps et antennes courtes.
25 % de drosophiles aux ailes écartées et antennes courtes.
Accidents méiotiques et évolution
Exercice : Accidents méiotiques et évolution
DOCUMENTS
Les bébés atteints du syndrome d’Edwards ont du mal à s’alimenter. Presque systématiquement on observe des malformations cardiaques mais aussi du squelette et des cartilages notamment au niveau du visage.
Tous ces troubles, et une sensibilité croissante aux infections sont des facteurs qui contribuent à la mort précoce de ces enfants.
Caryotype d’un fœtus atteint du syndrome d’Edwards
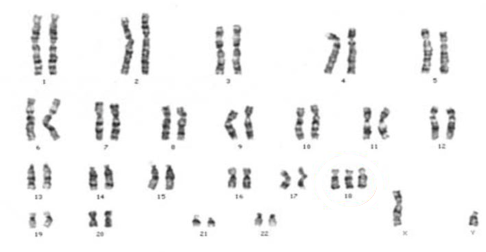
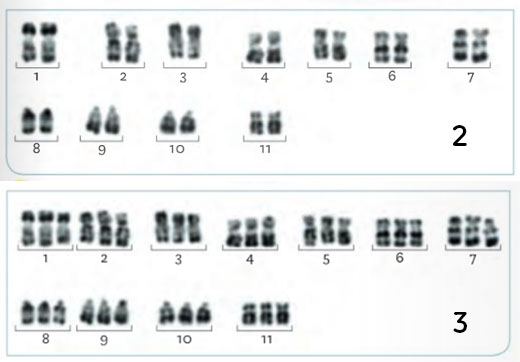
Caryotypes de la banane sauvage (2) et de la banane cultivée Cavendish (3)
ÉNONCÉ
Expliquer par des schémas les anomalies méiotiques aboutissant aux caryotypes 1 et 3, puis discuter de la notion « d’accident génétique » en lien avec l’évolution biologique.
Étude de cas : risque de transmission de la myopathie de Duchenne
Exercice : Étude de cas : risque de transmission de la myopathie de Duchenne
ÉNONCÉ
Un couple ayant déjà eu deux enfants, dont un enfant atteint de la dystrophie musculaire de Duchenne, (une maladie musculaire liée à une protéine, la dystrophine, non fonctionnelle) désire connaître le risque, pour leur enfant à naitre, d’être également touché par la maladie.
Identifier la cause génétique de la maladie, ainsi que le mode de transmission pour discuter du risque pour l’enfant à naître (III-3) d’être atteint.
DOCUMENT 1 : Arbre généalogique de l’enfant à naître
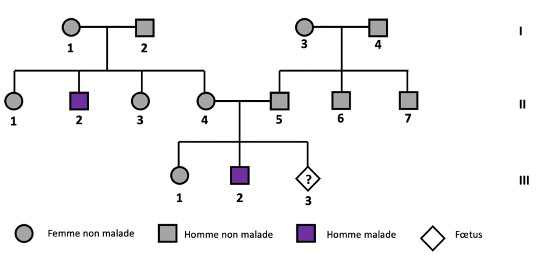
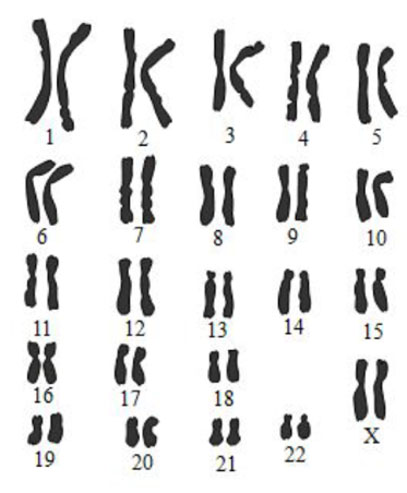
DOCUMENT 2 : Caryotype du fœtus III.3
Dans le cadre du dépistage prénatal proposé pendant la grossesse, le généticien qui suit la famille a proposé de réaliser un caryotype du fœtus. Les résultats sont présentés ci-dessous.
DOCUMENT 3 : Étude du gène de la dystrophine
Les séquences ci-dessous représentent la portion terminale du gène de la dystrophine, protéine des myocytes. Deux versions de ce gène sont représentées : l’allèle « sain » du gène, et l’allèle « malade » du gène.
Séquences brutes des nucléotides de la portion du gène :

Comparaison des deux séquences via Anagène :
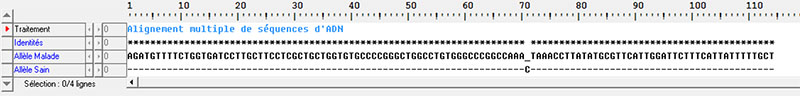
L’individu II-5 a été testé dans la famille étudiée, et n’est pas porteur de l’allèle malade.
Transferts horizontaux et endosymbiose
Exercice : Transferts horizontaux et endosymbiose
ÉNONCÉ
Énoncer l’hypothèse endosymbiotique de l’origine des mitochondries des cellules eucaryotes et dresser la liste des arguments présentés dans le document, en faveur de cette théorie.
Expliquer ensuite la différence notoire entre le nombre de gènes mitochondriaux et le nombre de gènes d’une bactérie actuelle.
DOCUMENT
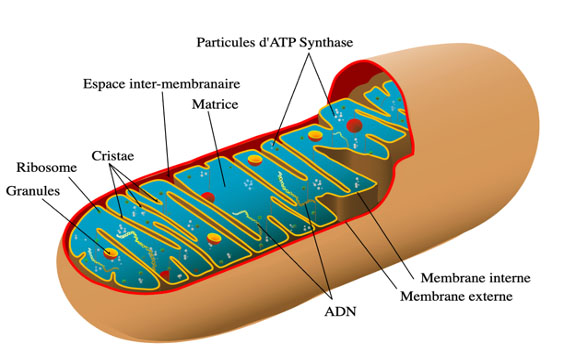
La mitochondrie est un organite retrouvé dans la plupart des cellules. Son diamètre varie généralement entre 0,75 et 3 µm tandis que sa forme générale et sa structure sont extrêmement variables. On en retrouve jusqu’à 2 000 par cellule. Elles permettent la production d’ATP et sont impliquées dans différents processus tels que la communication et la différenciation de la cellule, l’apoptose et la régulation du cycle cellulaire.
L’ADN des mitochondries est différent de celui du noyau.
La membrane mitochondriale externe contient l’ensemble de l’organite et a une épaisseur d’environ 6 à 7,5 nm.
Son rapport massique protéines/phospholipides est semblable à celui des membranes plasmiques des cellules d’eucaryotes. La membrane interne possède notamment un phospholipide, la cardiolipine, généralement caractéristique des membranes plasmiques bactériennes.
L’ADN mitochondrial (qui est le seul compartiment des cellules animales à posséder son propre ADN, en plus du noyau) est circulaire chez l’humain, et est composé de 16 569 paires de bases portant 37 gènes, codant des ARN messagers, des ARN de transfert, et des ARN ribosomiques. Ainsi la mitochondrie possède sa propre machinerie permettant la synthèse de protéines « sur place ». Les ribosomes des mitochondries sont petits et vulnérables aux antibiotiques.
Par comparaison, une bactérie possède toujours plusieurs centaines de gènes et le plus souvent plus de 1000.
La réplication de l’ADN mitochondrial n’est pas limitée à la phase S du cycle cellulaire. Indépendamment de la division cellulaire, la mitochondrie double sa masse puis se scinde en deux. Cette scission débute par l’apparition d’un sillon de division sur la membrane interne. Elle a lieu de multiples fois pendant toute l’interphase.
Mutations, clones cellulaires et génome de l'individu
Exercice : Mutations, clones cellulaires et génome de l’individu
ÉNONCÉ
On dit couramment que le génome est le même dans toutes les cellules d’un individu. Montrer en exploitant le document que ce n’est pas tout à fait exact et évaluer le nombre moyen de différences génétiques (mutations) possibles entre les génomes de 2 cellules d’un organisme de l’espèce E à la naissance.
DOCUMENT
On étudie une espèce animale E, présentée dans le tableau ci-dessous :
|
Durée du développement avant la naissance |
100 jours |
|
Nombre de mitoses avant la naissance |
50 |
|
Taille du génome |
16.109 nucléotides |
|
Taux de mutation |
15.10-9 nucléotide muté à chaque réplication du génome |
Quelques caractéristiques de l’espèce E
Survie, reproduction et équilibre de Hardy-Weinberg
Exercice : Survie, reproduction et équilibre de Hardy-Weinberg
DOCUMENT
On s’intéresse à une population composée de 5 000 individus d’espèce d’oiseaux exotiques (moitié mâles et moitié femelles). Il a été démontré que le volume du corps chez les mâles est principalement sous le contrôle d’une paire d’allèles : Vo+ et Vo. L’allèle dominant Vo+ permet un développement accru du corps.
Les mâles homozygotes récessifs Vo/Vo présentent un corps plus fluet. Or les mâles se battent aux beaux jours pour l’accès aux femelles, les mâles Vo/Vo ont donc un accès très limité à celles-ci, et l’on s’attendrait à ce que l’allèle Vo disparaisse au cours du temps.
Pourtant, les mâles homozygotes récessifs Vo/Vo représentent encore 12 % de la population mâle.
On dénombre 500 mâles Vo+/Vo+ dans la population étudiée.
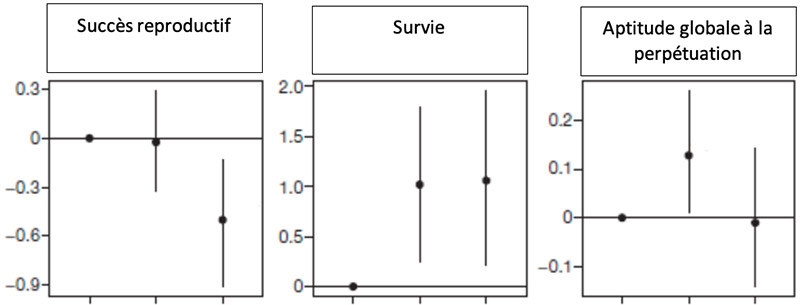
De la gauche vers la droite : oiseaux mâles de génotype Vo+/Vo+, Vo/Vo+, Vo/Vo et femelle

Des études concernant la survie en milieu naturel, et le succès reproductif ont été menées pour comparer les 3 génotypes des mâles.
Pour chaque graphique, la première colonne représente les mâles homozygotes dominants Vo+/Vo+ ; la deuxième, les mâles hétérozygotes Vo/Vo+ ; et la troisième, les homozygotes récessifs Vo/Vo.
Le « succès » des hétérozygotes dominants est donc pris comme valeur de référence (à 0) et le succès des autres lui est comparé (unités verticales arbitraires).
ÉNONCÉ
Montrer que la population mâle n’est pas à l’équilibre de Hardy-Weinberg concernant le gène étudié et proposer une explication à ce fait.
Étude d'un cas de spéciation
Exercice : Étude d’un cas de spéciation
DOCUMENT
L’aubépine est un arbre qui pousse partout en Amérique du Nord et produit un petit fruit (la cenelle), consommé par une larve de mouche parasite (genre Rhagoletis).
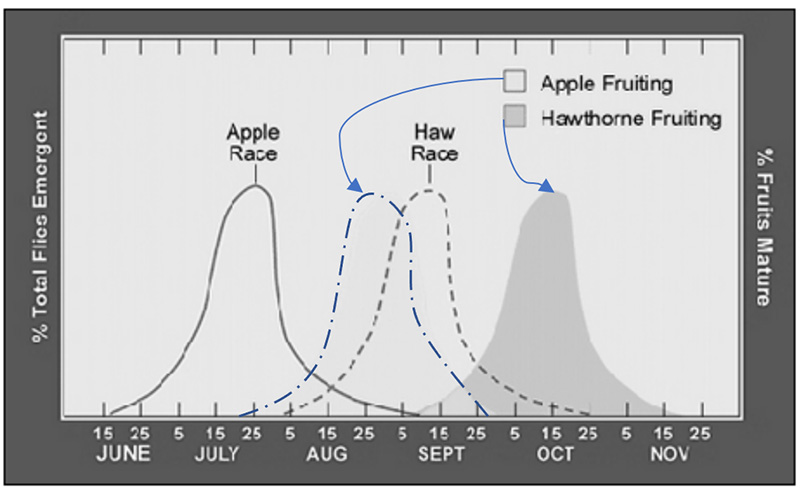
Lorsque les fruits mûrs tombent au sol les larves s’y enfoncent et passent l’hiver en vie ralentie (la durée de cette « vie ralentie » est codée génétiquement). La métamorphose de larve à adulte a lieu aux beaux jours et aboutit à l’apparition des adultes, qui coïncide avec la période de maturité des fruits de l’arbre hôte.
Étude comparée des deux populations de mouches
Au XIXe siècle, lors de la culture des premiers pommiers en Amérique du Nord, des mouches de l’aubépine ont commencé à parasiter les pommiers. Aujourd’hui, on observe que certaines mouches préfèrent pondre leurs œufs dans les cenelles alors que d’autres ont une forte préférence pour la ponte dans des pommes.
En laboratoire, les mouches du pommier et celles de l’aubépine s’hybrident facilement (descendance fertile) mais, dans la nature, les croisements entre les deux types sont peu fréquents. Pourtant, aucun caractère morphologique ou comportemental ne les différencie. Le taux d’hybridation entre la population de mouche se reproduisant dans la pomme et celle se reproduisant dans les cenelles n’est que de 4 % à 6 %.
Sortie du sol et métamorphose des mouches au cours de l’année
Maturité des pommes et cenelles

Étude des capacités de reconnaissance olfactive chez les mouches Rhagoletis
Si l’orientation des mouches parasites est visuelle, la reconnaissance spécifique de l’hôte peut être olfactive. Les adultes issus des larves qui se sont développées sur un hôte ont tendance à revenir sur ce même type d’hôte pour se reproduire du fait de la reconnaissance de composés volatils émis par l’espèce végétale. On suppose que l’attraction olfactive des mouches, qui dépend des propriétés des antennes, est d’origine génétique.