Catalyse enzymatique et apport de glucose sanguin



Catalyse enzymatique et apport de glucose sanguin
L’apport en sucre dans l’organisme se fait principalement par l’alimentation. L’amidon notamment se trouve dans de nombreux aliments. Il s’agit toutefois d’un glucide complexe, un polymère, qui a besoin d’être découpé en petits morceaux, les monomères de glucose, avant d’être distribués aux organes par la circulation sanguine. La réaction biochimique qui effectue ce découpage s’appelle l’hydrolyse.
L’hydrolyse requière cependant pour se réaliser in vivo un milieu d’une température nettement supérieure à celle du corps humain, qui se situe seulement autour de 37°C. Ce sont des protéines, les enzymes, qui agissent comme catalyseurs et permettent l’hydrolyse.
Les enzymes présentent par ailleurs une double spécificité :
– une spécificité de substrat : l’enzyme n’est capable d’agir que sur un seul substrat. Par exemple seule l’enzyme amylase est capable d’agir sur l’amidon,
– une spécificité d’action : l’enzyme n’est capable d’effectuer qu’une seule action. Par exemple, l’amylase est seulement capable de transformer l’amidon en glucoses.
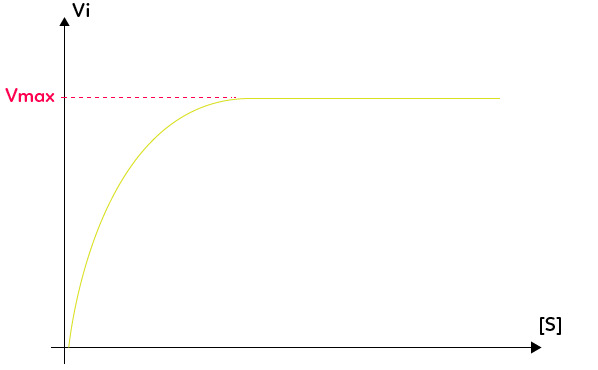
Si on suit expérimentalement la vitesse de la catalyse enzymatique en fonction de la concentration en substrat (on pourrait également suivre en fonction de la concentration en enzyme), on observe que lorsque la concentration de substrat dans le milieu augmente, alors la vitesse de la réaction augmente. Toutefois, cette augmentation de la vitesse de réaction n’est pas indéfinie, il y a un seuil, une vitesse maximale au-delà de laquelle on ne peut aller.
De ces observations et de celles de la double spécificité (de substrat et d’action), on a pu déduire qu’il existait une complémentarité spatiale entre l’enzyme et son substrat. Les deux molécules, du fait d’une forme tridimensionnelle complémentaire s’imbriquent l’une dans l’autre. On peut alors écrire la modélisation suivante :
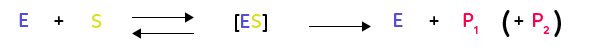
L’enzyme et le substrat forment effectivement un complexe enzyme-substrat (noté ci-dessus [ES]), qui permet à la catalyse d’avoir lieu. À la fin de la réaction, on retrouve alors l’enzyme intacte en plus du ou des produits. Il s’agit alors d’une autre particularité des réactions enzymatiques, les enzymes sont conservées intactes lors des réactions (elles sont ainsi bien des catalyseurs, ne réagissant pas directement), et peuvent donc être réutilisées.
On peut également modéliser une catalyse enzymatique de la façon suivante :
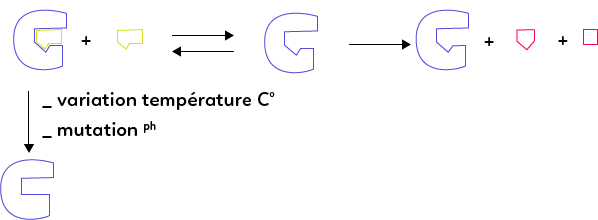
L’enzyme, représentée ici en bleu, possède une zone particulière composée de quelques acides aminés, appelée site actif de l’enzyme (qui a généralement la forme d’une poche) auprès duquel vient se fixer momentanément le substrat, et dans lequel la catalyse a lieu. On comprend donc l’importance première de la conformation (= forme) de l’enzyme. Cette forme en effet, obtenue lors de la maturation de l’enzyme, permet d’isoler et de protéger le site actif, de le conserver et de l’isoler du milieu dans lequel évolue l’enzyme.
Si l’enzyme venait à changer de conformation, alors le site actif pourrait ne plus être fonctionnel. Il existe un certain nombre de facteurs qui peuvent faire varier la conformation de l’enzyme, notamment :
– la variation de la température ;
– la variation du pH ;
– l’apparition d’une mutation génétique qui modifie la séquence d’ADN et à son tour éventuellement la séquence d’acides aminés correspondant à la protéine enzymatique, ce qui pourrait donc avoir un impact sur la conformation de l’enzyme.
Si les conditions du milieu dans lequel évolue l’enzyme sont modifiées (température ou pH), ou si une mutation génétique a lieu, alors l’enzyme se trouve partiellement ou totalement dénaturée, sa conformation change. Elle n’est alors plus capable d’accueillir le substrat et donc de catalyser la réaction.
Homéostasie et glycémie : la notion de variable régulée
Le milieu intérieur est l’ensemble des liquides extracellulaires de notre organisme : le sang, la lymphe et le milieu de vie des cellules.
L’homéostasie est la capacité de l’organisme à réguler un certain nombre de ses paramètres afin de les garder constants.
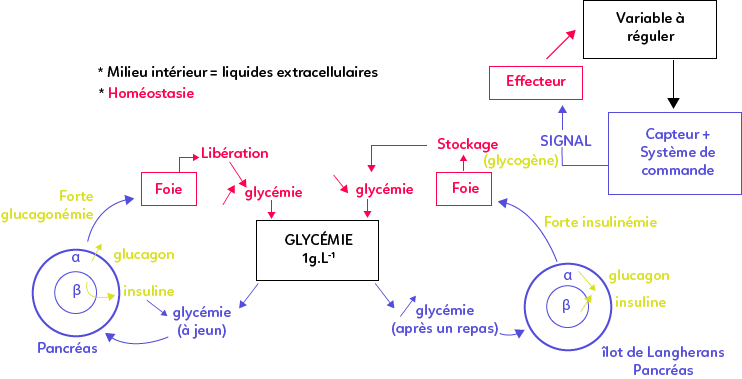
I. Homéostasie
Pour cette régulation, il existe une boucle applicable à l’ensemble des valeurs régulées dans le cadre de l’homéostasie. D’abord, il existe au sein de l’organisme des capteurs capables d’enregistrer la valeur qu’a la variable à un instant donnée. Ils transmettent l’information à un système de commande qui a la capacité de comparer la valeur enregistrée à une valeur que l’on appelle consigne. Si la valeur enregistrée est trop différente de la consigne, alors le système de commande envoie un signal (qui peut avoir différentes formes) à un organe ou à des cellules qui jouent le rôle d’effecteur. Les effecteurs sont alors capables d’avoir une action directe sur la variable à réguler, en l’augmentant ou la diminuant pour la rapprocher de la valeur consigne.
II. Application à la glycémie
Dans le cadre de la glycémie, une boucle de régulation existe. La valeur consigne est autour d’un gramme par litre (valeur moyenne de la glycémie chez l’être humain). Cette glycémie peut être régulée par l’intermédiaire du foie, qui joue le rôle d’effecteur. En effet, le foie est capable de stocker du glucose ou d’en libérer en fonction des besoins de l’organisme. Les besoins de nos cellules sont à peu près constants au cours de la journée, ils sont plus importants par exemple lorsque le cerveau fonctionne de façon importante ou lors d’un effort musculaire, et moins importants dans une situation de repos ou quand on dort. Les apports sont donc à peu près constants aux organes alors que notre consommation de sucre est discontinue étant donné qu’on en ingère seulement au moment des repas. Le foie joue donc le rôle d’intermédiaire.
D’autres organes sont capables de stocker du glucose : les muscles et le tissu adipeux. Néanmoins, ce sont plutôt des réserves « privées », c’est-à-dire que le stockage ne sert qu’aux cellules des muscles par exemple, il n’y a pas de distribution à d’autres organes, contrairement à ce que fait le foie.
Ce stockage se fait sous forme de glycogène, qui est un polymère de glucose, produit dans les cellules hépatiques ou musculaires lorsque la glycémie est haute. Quand la glycémie s’éloigne de la valeur consigne, repère, elle est mesurée, captée par les cellules du pancréas. C’est un organe à double fonction :
– exocrine (sécrète des substances destinées à être expulsées dans le milieu extérieur, au niveau de la peau, du tube digestif ou de l’arbre respiratoire),
– endocrine (sécrétant ses hormones directement dans le sang).
Ici, on s’intéresse à sa fonction endocrine car il sécrète des hormones, molécules chimiques libérées et circulant dans le sang, agissant à distance sur des cellules cibles qui possèdent des récepteurs hormonaux spécifiques à cette hormone et répondent à l’hormone en modifiant son fonctionnement. Deux hormones du pancréas nous intéressent : l’insuline et le glucagon.
Notre pancréas possède des îlots de Langherans au sein desquels on trouve des regroupements de cellules, des cellules beta capables de produire de l’insuline et des cellules alpha capables de produire du glucagon.
Lorsque le pancréas enregistre une hausse de la glycémie, il augmente sa production d’insuline et diminuer sa sécrétion de glucagon. On a donc une forte insulinémie (concentration d’insuline) dans le sang qui est un signal envoyé aux organes capables de stocker du glucose. À ce moment-là, les cellules hépatiques prélèvent du glucose dans le sang et la glycémie diminue grâce à ce stockage. On revient à une glycémie proche de la valeur consigne.
À l’inverse, quand on est en phase de glycémie basse (quand on est à jeun par exemple), il peut y avoir un risque pour certains de nos organes qui ont un besoin constant de glucose. On ré-augmente donc la glycémie. Le pancréas joue un rôle inverse : il diminue sa sécrétion d’insuline et augmente la sécrétion de glucagon. On a donc une forte glucagonémie dans le sang, signal pour le foie qui libère alors du glucose. On a alors une ré-augmentation de la glycémie et un retour à la valeur consigne.