Les étapes de la réaction immunitaire adaptative



Les étapes de la réaction immunitaire adaptative
I. Réaction immunitaire adaptative à médiation cellulaire
Qu’est-ce que la réaction immunitaire adaptative à médiation cellulaire ?
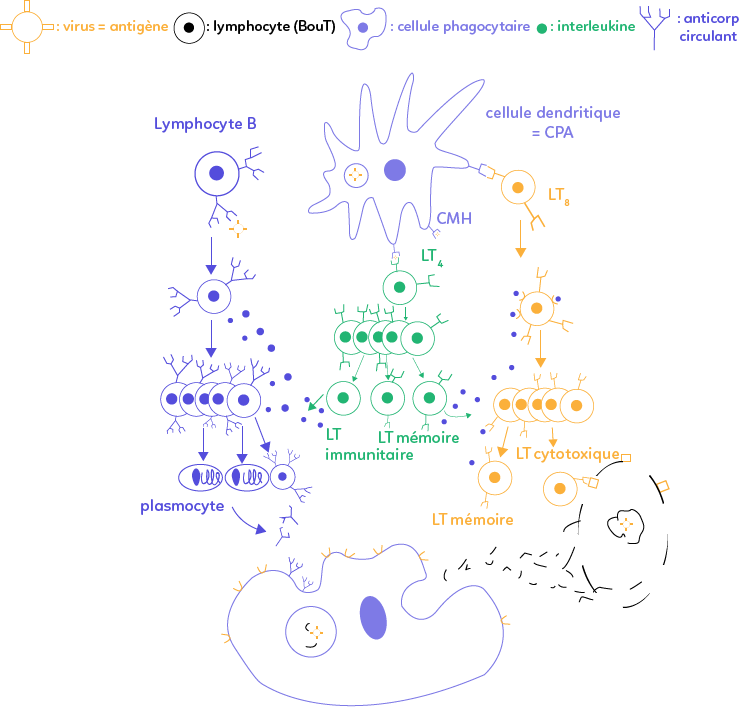
La réaction immunitaire adaptative débute avec les cellules dendritiques que l’on appelle cellules présentatrices d’antigène (CPA). Ces cellules peuvent phagocyter le virus et présentent ensuite à leur surface des morceaux du virus (des peptides viraux) associés au complexe majeur d’histocompatibilité (CMH). Elles activent alors les lymphocytes T4 et T8 qui sont incapables de reconnaître un antigène libre en solution. Ces cellules dendritiques ont donc une place maîtresse dans la réaction immunitaire adaptative (pour rappel, elles jouent également un rôle important dans la réaction immunitaire innée).
Une fois l’antigène présenté à la surface de ces cellules, les lymphocytes ayant sur leur membrane les récepteurs T complémentaires à l’antigène donné sont sélectionnés. Les lymphocytes T4 ainsi activés se multiplient puis se différencient en lymphocytes T auxiliaires (ou helper) ou deviennent des lymphocytes T mémoires de durée de vie plus longue.
À la surface des lymphocytes T8 sélectionnés apparaissent des récepteurs complémentaires de l’interleukine libérée dans le milieu par les lymphocytes T4 auxiliaires.
Une fois les interleukines logées dans les récepteurs, les lymphocytes T8 se multiplient et se différencient en lymphocytes T cytotoxiques qui en libérant les perforines entraînent la lyse d’une cellule infectée par un virus. Ainsi, un virus s’étant caché dans une cellule hôte est détruit par lyse cellulaire grâce aux lymphocytes T cytotoxiques dont la différenciation a été permise par les interleukines sécrétées par les LT auxiliaires, qui sont donc les cellules pivots de la réaction immunitaire. La destruction du virus par lyse cellulaire s’appelle la réaction immunitaire adaptative à médiation cellulaire.
II. Réaction immunitaire adaptative à médiation humorale
Qu’est-ce que la réaction immunitaire adaptative à médiation humorale ?
Cette dernière débute par les lymphocytes B, capables de reconnaître l’antigène sans entrer en contact avec la cellule dendritique grâce à leurs anticorps membranaires. Une fois ces derniers fixés à l’antigène, il apparaît à la surface des lymphocytes B des récepteurs à interleukines stimulés par l’interleukine libérée par les LT auxiliaires. Ils se multiplient puis se différencient alors en LB mémoires à durée de vie longue ou en plasmocytes, sécréteurs d’anticorps libres circulants qui forment ensuite des complexes immuns. Ces derniers s’agglutinent alors sur le virus et le neutralisent. Le virus est alors phagocyté et donc détruit par le macrophage.
Conclusion :
Il existe deux voies complémentaires de destruction du virus :
– la voie humorale par le biais des anticorps ;
– la voie cellulaire par le biais des lymphocytes T cytotoxiques qui lysent des cellules infectées.
On peut noter le rôle central des lymphocytes T auxiliaires qui stimulent la multiplication et la différenciation des deux voies.
L'immunité adaptative à médiation humorale
Quelle est la différence entre immunité innée et immunité adaptative ?
L’immunité adaptative s’oppose à l’immunité innée (réaction inflammatoire). On s’intéresse plus particulièrement à l’immunité adaptative à médiation humorale dont le médiateur, c’est-à-dire celui qui intervient majoritairement, est l’anticorps présent dans le sérum (dont l’ancienne appellation est « l’humeur »).
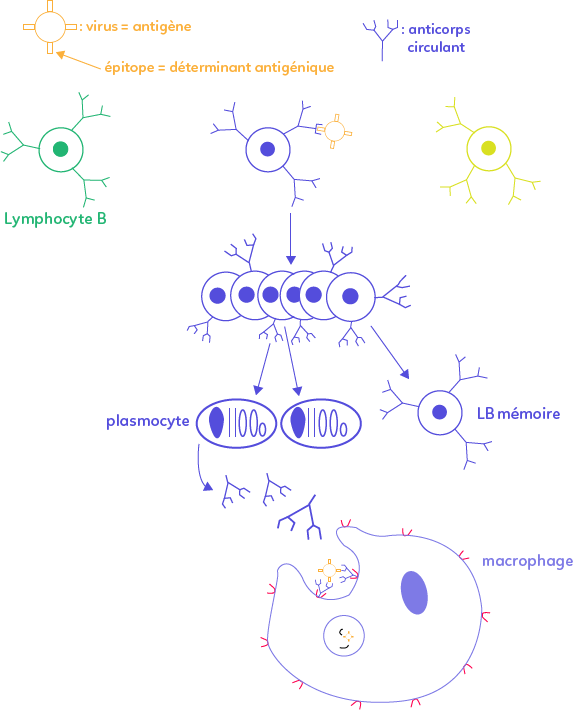
La réaction débute par l’introduction d’un antigène (une substance, un micro-organisme, une molécule) qui entraîne une réaction immunitaire.
Un virus porte souvent à sa surface des protéines (hémagglutinine dans le cas de la grippe) appelées épitopes ou déterminant antigénique. Ces dernières sont reconnues par des récepteurs membranaires ou par des anticorps membranaires présents à la surface des lymphocytes B. Au niveau des organes lymphoïdes secondaires préexistent à toute infection des clones de lymphocytes B, capables de reconnaître un épitope particulier.
Seuls certains clones sont capables de reconnaître un virus précis car ils ont, à l’extrémité de leurs anticorps membranaires, une partie hypervariable de forme complémentaire au déterminant antigénique du virus. Cette première phase correspond à la sélection clonale ou d’activation : le lymphocyte B n’avait encore jamais rencontré le virus, on parle de lymphocyte naïf.
À l’issue de cette activation, de nouvelles étapes se déclenchent.
Le lymphocyte B sélectionné se multiplie : c’est la phase de multiplication. Beaucoup de lymphocytes B se différencient ensuite en plasmocytes (l’autre partie se différenciant en lymphocytes B mémoires). Ce sont des cellules particulières de gros volume, avec un réticulum endoplasmique, un appareil de golgi très développé et de nombreuses cellules de sécrétion.
Les plasmocytes sécrètent des anticorps circulant librement dans le sérum (ils ne sont pas enchâssés dans la membrane). Les anticorps neutralisent ensuite le virus en s’y accrochant, ils forment alors un complexe immun qui empêche le virus d’infecter une cellule et de s’y multiplier.
Enfin, le complexe immun est phagocyté par une cellule phagocytaire (macrophage, cellule dendritique, granulocyte) créant alors un phagosome (une vésicule de digestion) qui fusionne avec d’autres vésicules contenant différentes enzymes digestives : le virus est ainsi détruit.
Le virus qui s’était introduit dans l’organisme a été détruit par immunité adaptative car elle est spécifique : on a sélectionné uniquement les clones de lymphocytes propres à ce virus.