Anomalies méiotiques aboutissant à des trisomies



Anomalies méiotiques aboutissant à des trisomies
Un individu trisomique est un individu qui possède dans toutes ses cellules somatiques (qui normalement sont diploïdes avec des chromosomes qui vont par paire) trois exemplaires d’un même chromosome. La trisomie la plus répandue est la trisomie 21.
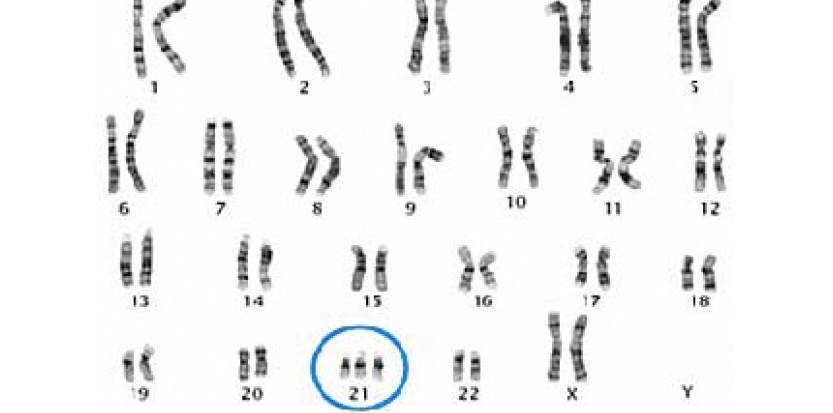
Sur le caryotype ci-dessus, on constate qu’au niveau de la paire 21, il y a, non pas une paire, mais un triplet de chromosomes donc trois chromosomes 21. Cette maladie est appelée le syndrome de Down. Elle est associée à divers dysfonctionnements au niveau physiologique et aussi à un phénotype macroscopique qui se reconnaît.
Certaines trisomies ne sont pas viables, c’est-à-dire qu’un individu humain ne peut pas vivre avec n’importe quel chromosome présent en trois exemplaires au lieu de deux.
D’autres trisomies sont viables, ce sont notamment les trisomies qui concernent les chromosomes sexuels. Par exemple, un caryotype humain « anormal » avec deux chromosomes X et un chromosome Y donc trois chromosomes sexuels au lieu de deux dans les cellules somatiques : c’est le syndrome de Klinefelter.
Il y a aussi le syndrome du triplo X, avec trois chromosomes X chez certaines femmes.
Comment un individu peut-il avoir trois chromosomes dans chacune de ces cellules ?
S’il y a une anomalie, cette anomalie a eu lieu chez les parents de cet individu (plus précisément chez un de ses deux parents) qui, lors de la formation des gamètes, a eu un dysfonctionnement méiotique produisant alors un ou des gamètes anormaux avec un chromosome en trop. L’individu est trisomique parce qu’un de ses parents a donné un gamète avec un chromosome en trop. Normalement chaque gamète possède un seul exemplaire pour chaque chromosome.
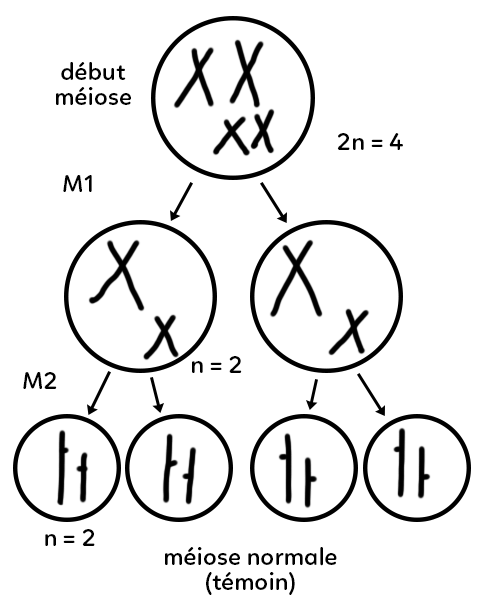
On représente schématiquement le déroulement d’une méiose normale (cas témoin). Une cellule avec deux paires de chromosomes homologues (qui pourrait être une cellule mère des gamètes) subit la méiose. Elle possède 2n = 4 chromosomes en début de méiose. En fin de méiose 1, les chromosomes homologues se sont séparés et ont migré chacun dans une des cellules filles. On obtient une cellule haploïde n = 2 qui porte un chromosome issu de chaque paire (un grand et un petit). Elle est haploïde et porte des chromosomes bichromatidiens. Au cours de la méiose 2, chaque chromosome voit ses deux chromatides se séparer de chaque côté de la cellule vers les pôles cellulaires.
En fin de méiose, on obtient des gamètes qui contiennent chacun n = 2 chromosomes et chacun de ces chromosomes porte une chromatide.
Dans certains cas de méiose, il y a une mauvaise séparation des chromosomes ou des chromatides. Cette séparation a lieu, normalement, en anaphase 1 pour la séparation des chromosomes homologues et en anaphase 2 pour la séparation des chromatides de chaque composant.
Anomalie en anaphase 1
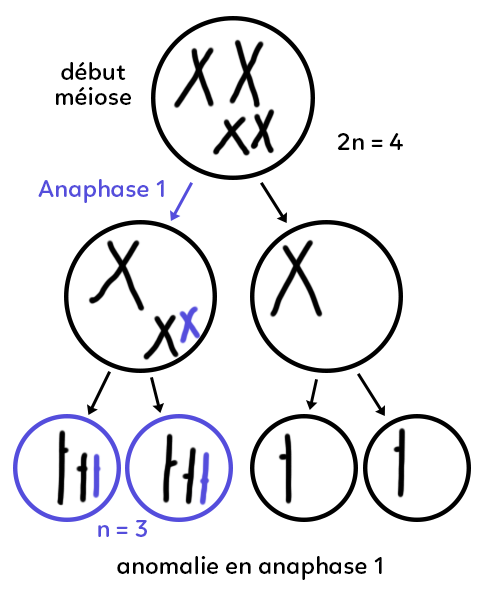
La cellule en début de méiose est comparable à celle de la première partie du schéma (2n = 4 chromosomes). S’il y a une mauvaise séparation des chromosomes au moment de l’anaphase 1. Normalement chacun des grands chromosomes devrait se retrouver dans les cellules filles. C’est le cas. En revanche, les deux petits chromosomes ont été vers la même cellule fille et il y en a donc un en trop (représenté en bleu) et un en moins dans l’autre. On parle de non-disjonction ou non-séparation des chromosomes homologues en anaphase 1.
Les chromatides de chacun de ces chromosomes vont se séparer au moment de l’anaphase 2. Si cette anaphase 2 se déroule normalement, on récupère deux gamètes (entourés en bleu) qui portent, non pas deux chromosomes comme c’est le cas dans la cellule témoin, mais un troisième chromosome : il y a deux exemplaires du petit chromosome au lieu d’un seul, donc « n = 3 ».
La présence de ces trois chromosomes ne veut pas dire que le gamète, lui, est trisomique. C’est l’enfant éventuellement issu de ces gamètes qui sera trisomique. Pour l’instant ce gamète a juste un chromosome en trop. On a aussi deux gamètes anormaux puisqu’il devrait avoir un petit chromosome et il n’y en a pas.
Quand l’anaphase 1 se passe mal et qu’il y a une anomalie en début de méiose, on obtient 4 gamètes anormaux : 2 qui possèdent un chromosome en trop et 2 qui possèdent un chromosome en moins.
Anomalie en anaphase 2
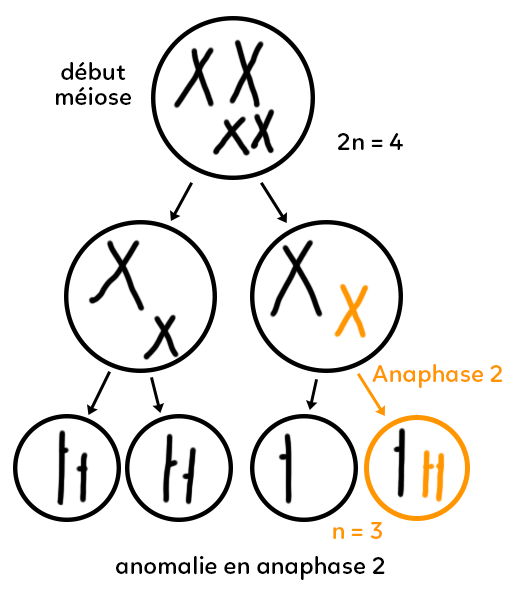
On part toujours d’une cellule avec 2n = 4 chromosomes avec une méiose 1 normale. Pour la cellule de la gauche, l’anaphase 2 se déroule normalement avec 2 gamètes qui portent chacun n = 2 chromosomes.
En revanche, l’anaphase 2 présente une anomalie. Les deux chromatides du petit chromosome ont migré du même côté. On parle de non-disjonction ou non-séparation des chromatides en anaphase 2.
Dans ce cas, on trouve un gamète (entouré en orange) qui possède trois chromosomes. Il a donc deux petits chromosomes au lieu d’un seul. Attention, ce gamète n’est pas trisomique, c’est la cellule œuf et donc le futur enfant qui pourrait hériter de ce gamète qui sera trisomique.
Conclusion
Dans ces trois cas, on a donc trois gamètes qui présentent un chromosome surnuméraire. Lorsqu’une une cellule présente un nombre anormal de chromosomes (en plus ou en moins) on dit que cette cellule est aneuploïde. C’est l’adjectif qui caractérise une structure qui n’a pas le bon nombre de chromosomes par rapport au cas sans anomalie.
Il faut imaginer que si une cellule œuf qui résulte de la fusion de deux gamètes hérite d’un gamète avec un chromosome surnuméraire, par exemple, si un gamète possède deux chromosomes 21 au lieu d’un seul, et d’un gamète normal, alors cette cellule œuf possèdera 3 chromosomes 21. Comme cette cellule œuf va se diviser par mitose, chaque cellule de l’embryon aura 3 chromosomes 21 au lieu de 2 donc l’individu sera trisomique.
Crossing-over inégal, duplication géniques et familles multigéniques
Les crossing-over sont des mécanismes normaux au cours de la prophase I de méiose. C’est le moment où les chromosomes sont appariés, ils sont tellement proches qu’il se produit parfois des enjambements (croisements précis à des endroits des chromatides) et des échanges de chromatides entre chromosomes homologues. Parfois les portions échangées ne sont pas équivalentes : elles ne portent pas forcément les mêmes gènes ou le même nombre de gènes.
I. Mécanisme de crossing-over inégal
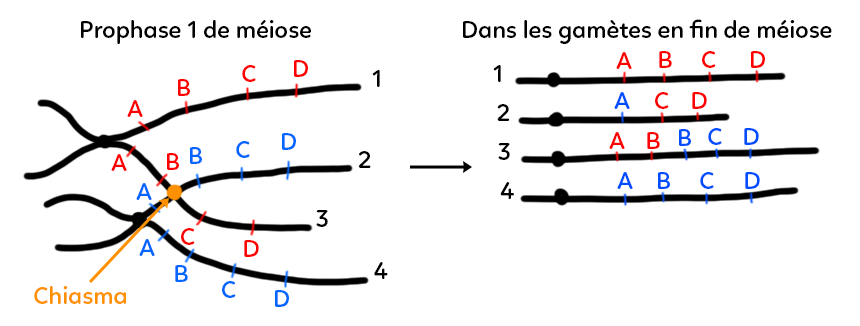
On a représenté une paire de chromosomes homologues appariés (prophase I). Ces chromosomes homologues sont dupliqués et portent chacun quatre gènes. Le chromosome numéro 1 a ses chromatides 1 et 3 qui portent chacune A, B, C et D (en rouge). Ce chromosome est apparié à son homologue dont on a représenté les gènes A, B, C et D (en bleu) sur les chromatides 2 et 4.
Le chiasma est un point d’enjambement au niveau duquel il peut se produire un échange entre les chromatides. Le chiasma se situe entre B et C sur le chromosome dont les gènes sont représentés en rouge alors qu’il se situe entre les gènes A et B sur le chromosome où les gènes sont représentés en bleu. Il n’y a donc pas d’équivalence pour les deux chromosomes homologues sur la position du chiasma. Lorsque le chiasma est mal situé (non placé au même endroit des chromosomes), s’il y a échange de portions de chromatides, ces portions ne vont pas être équivalentes. Sur le schéma, elles ne sont pas équivalentes dans leur longueur et donc dans les gènes qui vont être échangés.
Résultat de ce crossing-over inégal : un premier gamète porterait la chromatide 1 qui n’a pas été concernée par le crossing-over inégal (elle a toujours quatre gènes rouges A, B, C, D). En revanche, la chromatide 2 porte le gène A bleu et seulement C et D en rouge, elle a donc perdu le gène B. On parle d’une perte ou plus précisément d’une délétion génique. La chromatide 3 possède A et B en rouge et à nouveau B, C et D en bleu. Il y a eu un ajout car elle a deux fois le gène B. On parle d’une insertion génique. Puisqu’il y a deux fois le gène B dans ce gamète, on peut dire qu’il y a eu duplication (copie du gène B). La chromatide 4 n’a pas été concernée par le crossing-over et porte toujours A, B, C, D en bleu (c’est une chromatide inchangée). Pour les chromatides 2 et 3, c’est donc le résultat d’un crossing-over inégal : une délétion d’un gène entier et une insertion d’un gène entier.
II. Conséquences du crossing-over inégal
Ce crossing-over inégal a abouti à une duplication du gène B. Ainsi, dans les générations qui vont suivre, imaginons que le gamète qui contenait la chromatide numéro 3 soit utilisé et donne naissance à un individu qui vit normalement. Supposons que la duplication de ce gène B n’apporte pas de handicap mettant en péril la survie, soit de la cellule œuf, soit de l’individu. Imaginons que ce gamète donne naissance à un nouvel individu qui va lui-même transmettre son patrimoine génétique à sa descendance, etc. Au fil des générations, que va-t-il se passer ?
Nous avons maintenant dans cette lignée deux versions du gène B. On sait que l’ADN subit des mutations aléatoires et imprévisibles au cours du temps. Donc plus il y a de générations, plus les deux versions du gène B vont subir des mutations. Cependant, comme ces mutations sont aléatoires et qu’elles se produisent sur l’ensemble des gènes, les mutations vont être différentes pour les deux versions du gène B possédées par cette lignée. Il y a donc des mutations distinctes qui vont finalement donner naissance à deux versions légèrement différentes (mais de plus en plus différentes) du gène B. On peut les appeler B et B’. Chaque version de ce gène va évoluer de son côté, c’est-à-dire subir ses propres mutations qui, au fil du temps, vont transformer très légèrement ce gène B.
III. Familles multigéniques
Sur ce chromosome, on a deux gènes identiques au départ, qui sont devenus B et B’. Auparavant, ils étaient côte à côte dans le génome. On peut supposer que les gènes B vont soit rester proches, soit éventuellement être séparés l’un de l’autre (il existe parfois certains mécanismes génétiques qui déplacent des portions de chromosomes dans le génome).
Ces deux gènes vont avoir des séquences nucléotidiques très ressemblantes. On peut les comparer avec des logiciels comme Anagène ou GénieGen. Si on comparait B et B’, on trouverait un fort pourcentage de nucléotides en commun entre la séquence B et la séquence B’.
Les rôles (fonction au niveau moléculaire et au niveau de l’organisme de ces gènes) vont rester comparables. Si les gènes B et B’ ont des séquences nucléotidiques proches, alors les protéines seront elles aussi à priori ressemblantes et pourraient jouer des rôles comparables dans l’organisme. Mais, il arrive parfois qu’une simple mutation puisse changer complètement la forme ou le rôle de la protéine codée.
Ces gènes B et B’ qui évoluent distinctement l’un de l’autre forment alors une famille multigénique. Il s’agit d’un ensemble de deux (ou plus) de gènes qui ont une origine commune. Dans l’exemple, cette origine est due à un crossing-over inégal qui a donné naissance sur un chromosome (le chromosome numéro 3) à deux versions du gène B. Ces versions évoluent ensuite séparément, subissent leurs propres mutations mais constituent une famille multigénique dans le sens où elles vont se ressembler au niveau nucléotidique.
On peut prendre l’exemple des opsines qui sont des protéines permettant de capter la lumière dans la rétine. Elles font partie des photorécepteurs. Il en existe trois types chez l’homme : l’opsine S, l’opsine M et l’opsine L. Ces trois protéines sont codées par des gènes qu’on considère faisant partie d’une famille multigénique, c’est-à-dire qu’ils descendent tous les trois d’un unique gène ancestral.