Transferts horizontaux chez les bactéries



Transferts horizontaux chez les bactéries
On connait déjà le transfert vertical d’une génération à l’autre via la reproduction sexuée par exemple, mais il existe aussi des transferts d’information génétique horizontaux entre individus, par exemple, d’espèces différentes. Ces transferts horizontaux sont bien connus et ont été étudiés notamment chez les organismes bactériens, les eubactéries et les archées.
I. Qu’est-ce qu’une bactérie ?
Les bactéries sont des organismes unicellulaires anciennement appelés procaryotes qui font entre 5 et 10 µm (observables au microscope optique mais dont les détails seront vus avec des microscopes électroniques en transmission ou à balayage).
II. Quelles sont les formes des bactéries ?
Les bactéries peuvent avoir différentes formes : bâtonnet, coque, spiralée.
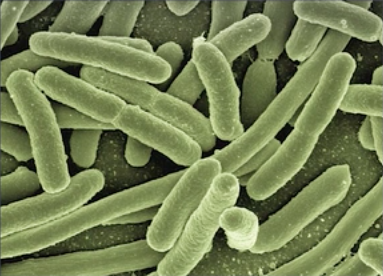
– Forme en bâtonnet, comme Escherichia coli qui constitue 80 % de la flore intestinale. Sur cette vue elle est observée au microscope électronique à balayage. Elle a une forme en bâtonnet, on parle de bacille. D’ailleurs bactérie en grec veut dire « bâtonnet ».
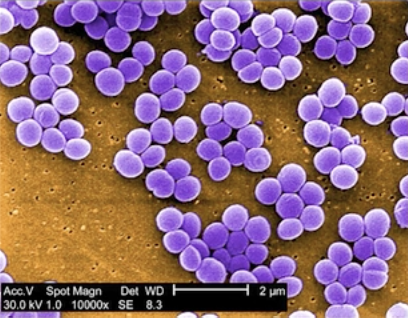
– Forme en coque, comme le staphylocoque doré (staphylocoque aureus) qui est une bactérie très connue, plutôt pathogène. Elle est vue en microscopie électronique à balayage et on remarque qu’elle se dispose en chaînettes. Il existe d’autres formes comme les coccobacilles, un compromis entre coque et bâtonnet.
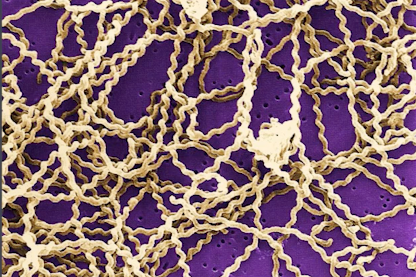
– Formes spiralées : les bactéries peuvent aussi avoir une forme spiralée comme sur cette image, bifides, isolées, en chaînettes ou en amas. Elles occupent de nombreux écosystèmes, jusqu’aux plus extrêmes. Les archées sont des bactéries souvent dites extrêmophiles, capables de vivre à des températures, des pressions et des conditions hygrométriques très particulières.
Ces bactéries ont la capacité de transférer leur matériel génétique de diverses façons, ce qui peut leur donner, par exemple, une certaine résistance aux antibiotiques. Ces types de transferts sont la conjugaison bactérienne, la transformation bactérienne et la transduction via des phages.
III. Comment se fait la conjugaison bactérienne ?
Historiquement, la conjugaison bactérienne a été mise en évidence par Lederberg et Tatum en 1946. Le principe était le suivant : Tatum et Lederberg ont créé des bactéries mutantes, dites auxotrophes pour certaines molécules organiques.
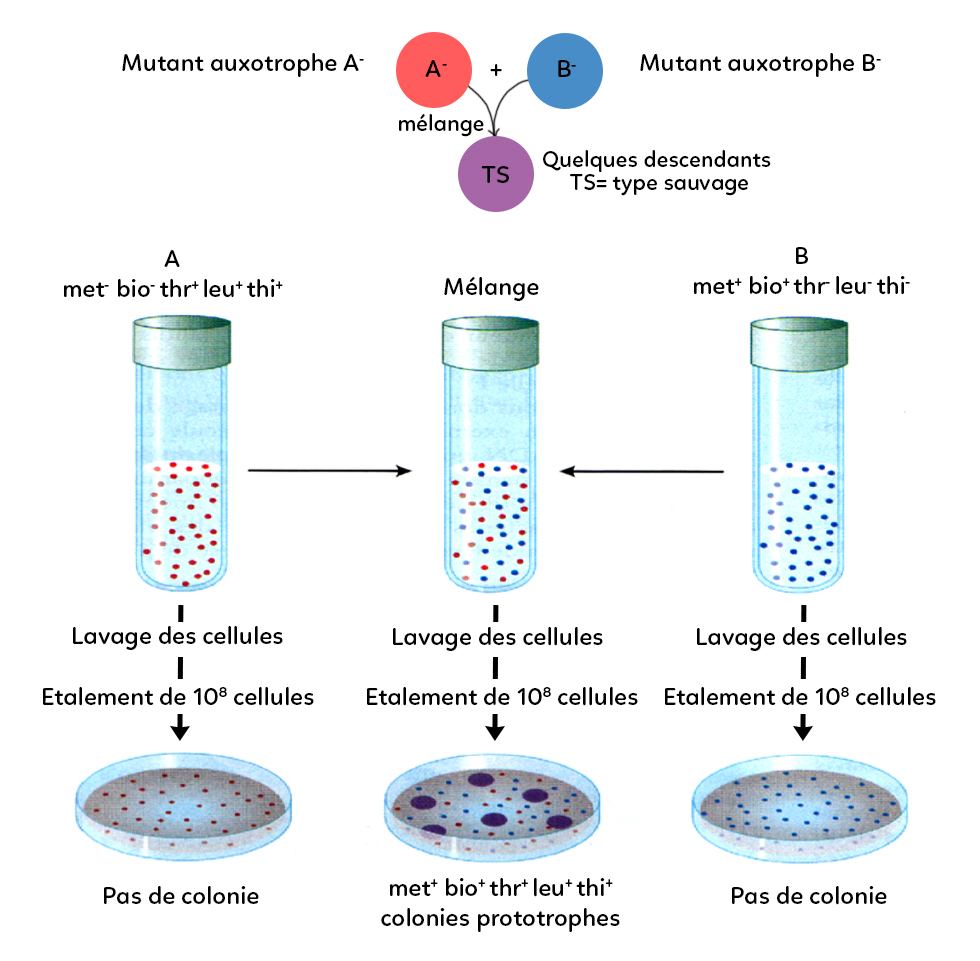
Dans ce schéma, le mutant A auxotrophe A- ne peut pas se développer si on ne lui donne pas de méthionine et de biotine. Le mutant auxotrophe B- ne peut pas se développer si on ne lui donne pas de thréonine, de leucine et de thiamine.
Si on met ces mutants A- et B- dans des milieux dits minimum, (sans méthionine, sans biotine, sans thréonine, sans leucine et sans thiamine), ils ne peuvent pas se développer.
Ensuite Tatum et Lederberg ont pris un tube à essai et y ont mélangé les mutants auxotrophes A et B puis les ont déposés sur une boîte de pétri en milieu minimum. Ils ont ensuite observé l’apparition de colonies qui arrivent à se développer bien qu’elles soient issues du mutant qui n’aurait pas pu se développer normalement. Ils en ont déduit que les mutants A et B ont pu échanger leur matériel génétique puisqu’ils sont complémentaires d’un point de vue génotypique. Cet exemple a permis de mettre en évidence la notion de conjugaison bactérienne.
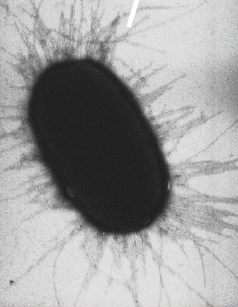
La conjugaison bactérienne dépend de la mise en place d’un petit pont cytoplasmique, un pilus (visible sur cette photo), uniquement fabriqué lorsque la bactérie dispose d’un plasmide F (fertilité). Cette bactérie qui dispose d’un plasmide (petit ADN circulaire), peut l’injecter dans une bactérie receveuse. Elles s’échangent ainsi de l’information. Lederberg et Tatum ne se sont pas arrêtés là, ils ont mis également en évidence que c’était par un contact bactérie-bactérie que la conjugaison bactérienne pouvait se faire. Grâce à un protocole avec un tube en U et une barrière filtrante, ils ont mis d’un côte les auxotrophes A-, de l’autre les auxotrophes B-. Les mutants ne pouvaient pas rentrer en contact du fait de la barrière qui ne laissait passer que des molécules. Aucune conjugaison bactérienne n’était observée, ce qui prouva que pour que les bactéries s’échangent de l’information, elles ont besoin de créer ce pont cytoplasmique, d’où un contact.
IV. Comment se fait la transformation bactérienne ?
La transformation bactérienne a été découverte en 1928, grâce aux expériences historiques de Griffith sur des pneumocoques, dont le principe est représenté ici.
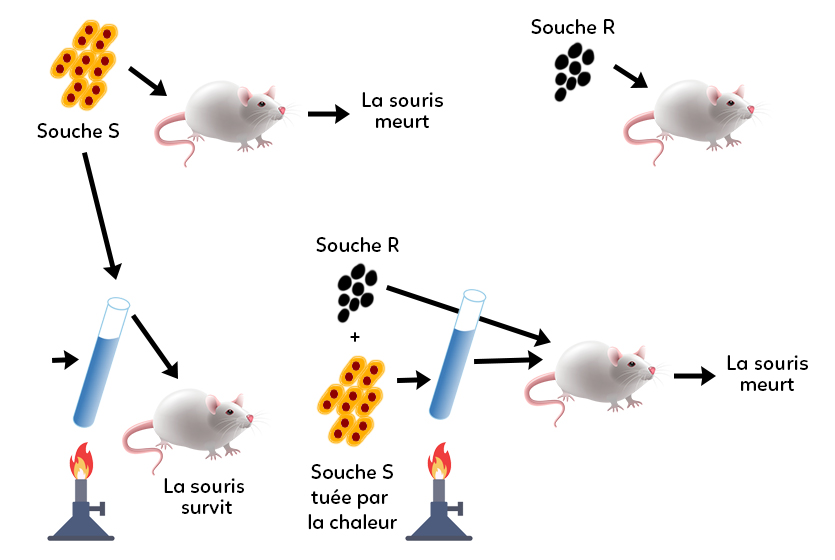
Il a travaillé sur un pneumocoque virulent dont il existait deux souches : une souche R (non virulente) et une souche S (virulente). Lorsqu’on injecte le pneumocoque S dans une souris, elle meurt. Lorsqu’on injecte le pneumocoque R dans une souris, elle survit.
Griffith a proposé de prendre une souche virulente, de la détruire et ensuite de la mettre en contact d’une souche non virulente, puis d’injecter l’ensemble dans une souris. Il constate que la souris meurt. On en déduit que la souche virulente morte a transmis son ADN. Griffith a parlé de « facteur transformant » à la souche non virulente, alors devenue virulente. C’est le principe de la transformation bactérienne. En 1928, la notion d’ADN n’existait pas : Griffith a juste mis en évidence qu’une bactérie pouvait devenir virulente par transformation en récupérant, par des bactéries virulentes, le facteur qui permettait d’être virulent. Ce facteur était de l’ADN. Cette récupération de molécules d’ADN se trouvant dans le milieu environnant se fait par des pores au niveau de la paroi et de la membrane de la bactérie.
V. Comment se fait la transduction ?
La transduction via des phages, étudiée par Hershey et Chase, a permis de comprendre grâce que la molécule appelée « facteur transformant » par Griffith était de l’ADN. Le phage, qui est un virus, va injecter son ADN ou son ARN dans une bactérie hôte et s’incorporer dans l’ADN de cette bactérie hôte en la modifiant. Si elle se multiplie, elle va conserver cet ADN viral. Si elle est parasitée par une autre espèce de phage, on va assister ainsi à des transferts horizontaux entre différentes espèces, phages et bactéries.
Conclusion
Avec ces trois types de transferts, on comprend que les bactéries, (archées, eubactéries) sont capables de transférer de l’ADN non pas uniquement par transfert vertical mais aussi par transfert horizontal entre espèces différentes, même avec des virus ce qui est une formidable façon de diversifier le vivant.
Théorie endosymbiotique
I. Des constats à l’origine de la théorie
Quelle est la théorie endosymbiotique ?
En observant une cellule végétale au MO, on remarque la présence de chloroplastes. On peut également observer d’autres organites cytoplasmiques que sont les mitochondries, présentes chez les organismes de type animaux, champignons et chez les protistes. Les chloroplastes et les mitochondries sont des organites présents dans le cytoplasme des cellules actuelles. Elles montrent des particularités structurales, notamment une double membrane, la présence de petits ADN circulaires et une certaine autonomie dans la réplication, dans la multiplication.
En observant en parallèle le fonctionnement de certaines cellules, on remarque des processus d’endocytoses : une cellule est capable de déformer sa membrane pour capturer un organisme qui était extracellulaire. Quand on fait le lien entre les organites actuels présents au sein des cytoplasmes des cellules végétales et animales, et la possibilité qu’ont les cellules de capturer du matériel extracellulaire, on construit la théorie endosymbiotique. « Endosymbiose » sous-entend que des cellules auraient capturé d’autres cellules par endocytose avec lesquelles elles auraient vécu en symbiose : cette relation aurait abouti à l’existence de ces organites semi-autonomes.
II. Étapes expliquant la théorie endosymbiotique
Comment se passe l’endosymbiose ?
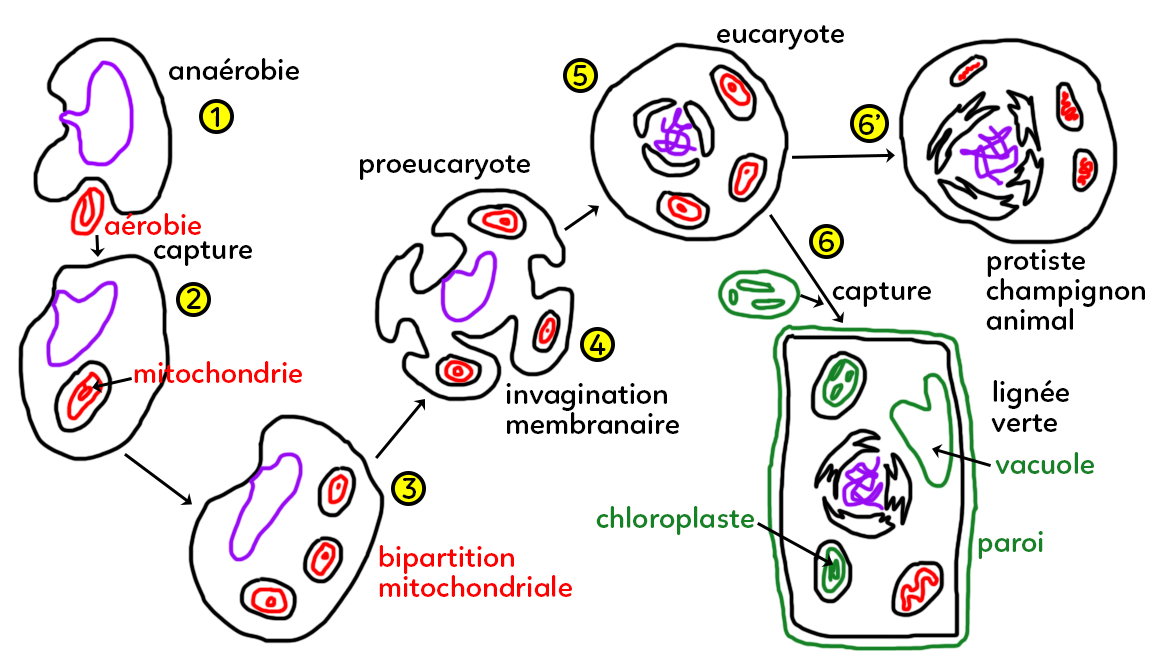
A. Étapes 1 à 2 : les premières mitochondries
On commence à plus de 2,4 Ga (milliards d’années), sachant que la vie sur Terre s’est développée il y a 3,4 Ga, avec des organismes unicellulaires chlorophylliens exclusivement aquatiques. On suppose donc qu’un organisme procaryote (sans noyau ; on parle maintenant de bactéries ou d’archées) anaérobie (avec un métabolisme qui n’utilise pas le dioxygène) aurait capturé un petit organisme aérobie (qui utilise le dioxygène, capable de faire le métabolisme respiratoire) par une déformation membranaire et l’aurait intégré dans son cytoplasme, d’où le fait que les organites possèdent une double membrane (la leur et celle de la vésicule d’endocytose). Cet organisme qui était anaérobie, grâce à cette capture, serait devenu à son tour aérobie. Ces organismes capturés seraient les premières mitochondries.
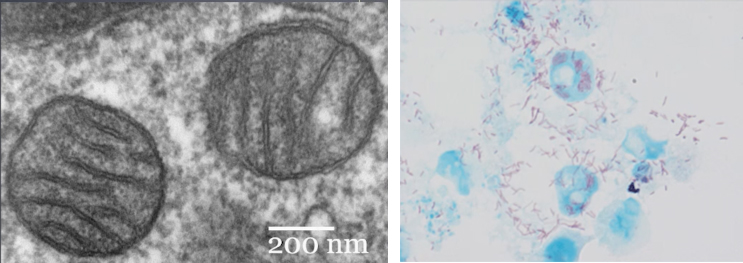
Lorsqu’on étudie la membrane externe, la membrane interne et le contenu génétique d’une mitochondrie actuelle, on trouve une grande ressemblance entre cette mitochondrie (photo haut) et les actuelles rickettsies (bactéries aérobies exclusives, photo bas). Par principe d’actualisme, on peut supposer que des bactéries qui ressemblaient aux rickettsies actuelles il y a 2,4 Ga, auraient été capturées pour former un procaryote aérobie (étape 2 du schéma).
B. Étapes 2 à 3 : la bipartition mitochondriale
Ce procaryote qui possède maintenant un organite de type mitochondrial voit ses mitochondries continuer de se multiplier (sans que la cellule ne se multiplie) par bipartition mitochondriale. On fait une analogie entre cette capacité qu’ont les mitochondries de se multiplier toutes seules avec la scissiparité observée chez les procaryotes.
C. Étapes 3 à 5 : l’apparition des eucaryotes
On est toujours chez un organisme procaryote sans noyau et au cours du temps, cet organisme aurait procédé à des invaginations de la membrane externe (étape 4) ce qui aurait abouti à un proeucaryote qui deviendra un eucaryote au sens strict (étape 5). Les eucaryotes (« Eu » = vrai ; « Caryote » = noyau) ont un vrai noyau constitué d’une enveloppe nucléaire (deux membranes accolées). À l’étape 5, on a donc un eucaryote aérobie avec ses mitochondries.
D. Étapes 5 à 6 ou 6’ : l’aboutissement aux lignées actuelles
Cet eucaryote aérobie va continuer de se développer. Il aboutit selon l’étape 6’ à un organisme unicellulaire ou pluricellulaire (champignons, animaux) fait d’un noyau et de plusieurs mitochondries.
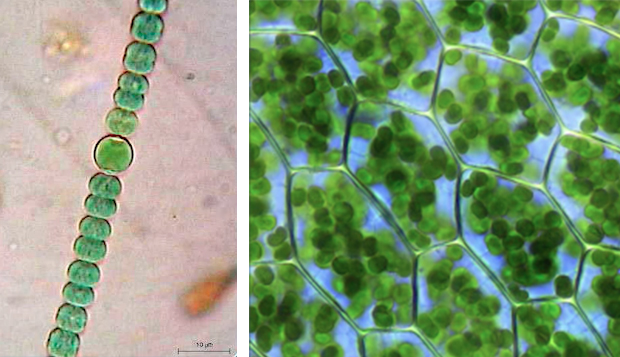
Mais à l’étape 5, cet eucaryote peut aussi avoir capturé au cours de son évolution une bactérie de type cyanobactérie actuelle visible ci-dessus à gauche, capable de réaliser le métabolisme photosynthétique avec des thylacoïdes et un petit chromosome circulaire. Capturé, il devient un nouvel organite : le chloroplaste (à droite), constitué de deux membranes, une externe et une interne. On aboutit ainsi à la lignée verte actuelle qui se caractérise par la présence de chloroplastes permettant un métabolisme photosynthétique, la présence d’une vacuole et la matrice extracellulaire de type paroi.
Conclusion
La théorie endosymbiotique permet d’expliquer l’existence d’organismes de la lignée verte avec des chloroplastes et des mitochondries, et pour les autres organismes (champignons, animaux), d’organismes avec des mitochondries exclusivement.
On a utilisé le terme semi-autonome : la multiplication de ces organites dépend en partie de la cellule totale. En effet, une partie des gènes de l’organisme ancestral qui a été capturé est passé dans le génome de l’organisme hôte, de sorte que les chloroplastes et les mitochondries dépendent en partie dans leur réplication, dans l’expression de leur génome, des gènes inclus dans le génome hôte, et donc de l’hôte. Il y a une sorte d’asservissement des organites au génome de la cellule totale.